Operationalizing replicator dynamics and partitioning fitness functions
February 25, 2015 18 Comments
As you know, dear regular reader, I have a rather uneasy relationship with reductionism, especially when doing mathematical modeling in biology. In mathematical oncology, for example, it seems that there is a hope that through our models we can bring a more rigorous mechanistic understanding of cancer, but at the same time there is the joke that given almost any microscopic mechanism there is an experimental paper in the oncology literature supporting it and another to contradict it. With such a tenuous and shaky web of beliefs justifying (or just hinting towards) our nearly arbitrary microdynamical assumptions, it seems unreasonable to ground our models in reductionist stories. At such a time of ontological crisis, I have an instinct to turn — much like many physicists did during a similar crisis at the start of the 20th century in their discipline — to operationalism. Let us build a convincing mathematical theory of cancer in the petri dish with as few considerations of things we can’t reliably measure and then see where to go from there. To give another analogy to physics in the late 1800s, let us work towards a thermodynamics of cancer and worry about its many possible statistical mechanics later.
This is especially important in applications of evolutionary game theory where assumptions abound. These assumptions aren’t just about modeling details like the treatments of space and stochasticity or approximations to them but about if there is even a game taking place or what would constitute a game-like interaction. However, to work toward an operationalist theory of games, we need experiments that beg for EGT explanations. There is a recent history of these sort of experiments in viruses and microbes (Lenski & Velicer, 2001; Crespi, 2001; Velicer, 2003; West et al., 2007; Ribeck & Lenski, 2014), slime molds (Strassmann & Queller, 2011) and yeast (Gore et al., 2009; Sanchez & Gore, 2013), but the start of these experiments in oncology by Archetti et al. (2015) is current events[1]. In the weeks since that paper, I’ve had a very useful reading group and fruitful discussions with Robert Vander Velde and Julian Xue about the experimental aspects of this work. This Monday, I spent most of the afternoon discussing similar experiments with Robert Noble who is visiting Moffitt from Montpellier this week.
In this post, I want to unlock some of this discussion from the confines of private emails and coffee chats. In particular, I will share my theorist’s cartoon understanding of the experiments in Archetti et al. (2015) and how they can help us build an operationalist approach to EGT but how they are not (yet) sufficient to demonstrate the authors’ central claim that neuroendocrine pancreatic cancer dynamics involve a public good.
First, I want to describe my understanding of the experiments that Archetti et al. (2015) carried out to make the right hand of figures 1B and figure 3C in their paper. I will sweep some details under the rug in hopes of being more general, but I think the main outline is accurate. If you are an experimentalist, or work with experimentalists, then please leave a comment to correct my oversights.
From conditioned media to mixed media
For the conditioned medium experiments in figure 1B, a petri-dish (or a well in a plate, or some such container) is filled with a liquid containing some food (the medium) and then cells of type X from some established culture are introduced onto this medium. They spend some fixed amount of time in that dish, consuming a portion of whatever is in the medium[2] and potentially releasing their own chemicals into the liquid. After a relatively short amount of time — something on the order of the cell cycle — the cells are separated from the medium, usually by running the mixture through a filter that is fine enough to catch cells but let macro-molecules through[3]. This gives us the post-X medium in a flask, which we pour out into a fresh petri-dish and now introduce a known number of cells of type Y onto it. Again, we grow this mixture for a fixed amount of time on the order of a cell cycle and then count how many cells there are. The ratio of cells that we introduced to the cells we count at the end is then the fitness of Y grown in medium conditioned by X.
Archetti et al. (2015) run the four combinations with each of X and Y being cells that are producers or nonproducers of IGF-II[4]. They observe that producers grow at about the same rate in media conditioned by producers or by non-producers, but non-producers grow much faster in media conditioned by non-producers than by producers. Further, non-producers grow slower than producers in media condition by non-producers, and faster than producers in media conditioned by producers. This is the sort of profile we would want to see when looking for a game-like dynamic.
Here, there seems to be an obvious extension that Archetti et al. (2015) do not consider, but that I would like to see run[5] — let’s call it the mixed media experiment. In particular, it seems like mixing beakers of liquid is a very easy experimental procedure, even one that a theorist like me could do. So why not take a beaker of conditioned medium from cells of type A and another from cells of type B and mix them in some proportion p. Now we can grow cells of type Y on this mixed medium and the ratio of cells that we introduced to the cells we count at the end will give us the fitness of Y as a function of the proportion p of type A medium in the A-B mix.
Mixed population and replating
Instead of my proposed mixed medium experiments, for figure 3C Archetti et al. (2015) ran a mixed populations experiment. Here, a petri-dish (or a well in a plate, etc) with a specific mix of medium is populated with a specific number of cells of type A and a specific number of cells of type B such that the type A cells form a specific desired initial proportion p of the cells. The cells are allowed to grow (maybe with a couple of replatings, it is not obvious to me from the description in the paper) for a few days and are then counted through flow cytometry[6] to figure out the new proportion of type A cells. The difference between the initial and final proportion is then plotted, and the experiment is repeated for different initial proportions p and — in the case of Archetti et al. (2015) — for different initial mixes of media.
Archetti et al. (2015) showed that as the concentration of serum in the medium is increased from 1% to 10%, producers become relatively less fit. This meshes nicely with the observations that more austere environments tend to encourage cooperation, but the authors prefer to interpret the increase in serum as increasing the cost/benefit ratio of IGF-II. More importantly, they show that for some concentrations of serum (5% and 6% are most dramatic) there is a clearly non-linear response in the relative fitness of producer cells. In particular, for high or low initial proportion p of producer cells non-producers fare better, but for medium initial proportions p the producers fare better. This allows the two cell types to exist in a polyclonal equilibrium, something that is not theoretically possible for replicator dynamics of a linear public good in a well-mixed population[7]. Unfortunately, the authors do not go beyond the typical approach in EGT of looking at qualitative correspondence and do not bother to fit specific fitness functions or replicator dynamics to their data[8].
Finally in figure 2, Archetti et al. (2015) consider long-term dynamics by running their experiment for 6 weeks. Although they don’t give great detail on their experiment, I will assume that it follows a standard replating procedure. From how often their data points are presented, it looks like they replated every seven days[9]. Standard replating proceeds as follows (as I discussed in my earlier post): (1) fill a new petri dish with a mix of nutritional medium[10] and put a known number N of two different cell types A and B on the medium (on the very first plate we will also know the proportion of A and B in the mixture); (2) let them grow for a fixed amount of time; (3) Randomly sample N cells from the grown population to repeat and seed (1) and then take another sample to run through flow cytometry to estimate the growth in A and B cells. This gives us a time series of the cells evolving that we can track indefinitely; Lenski has been following a similar procedure for decades with the E.coli long-term evolution experiment (Lenski et al., 1991; Wiser et al., 2013; Ribeck & Lenski, 2014).
Giving operationalist meaning to replicator dynamics
The point of philosophical schools like operationalism or instrumentalism — or maybe (analytic) philosophy more generally — is to make sure that we understand what we mean by the words we use. They propose we do this by making sure that the words we use are restricted to specific measurements in a fixed experimental protocol, instead of to some slippery reductionist ontology[11]. In the case of evolutionary game theory, the central word we need to define is ‘fitness’.
Let’s look at the structure of the replating experiment. If we originally had pN cells of type A and (1 – p)N cells of type B and they grew on our plate to numbers 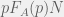



You might recognize the above as the discrete-time replicator equation[12] where 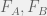
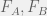
Somehow, this might seem unsatisfying in that we just named something but aren’t testing anything. This is where we can begin to twist our experimental knobs a bit to start understanding our system. For example, consider the mixed media experiments described above, let us use them to define another notion of ‘fitness’: experimental medium-mediated fitness. When we grow cells X on media conditioned by a proportion p of type A conditioned media in the A-B mix then we can call the corresponding ratio between initial and final cells as 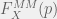
Now, if we compare the experimental medium-mediated fitness to the experimental fitness 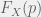

In general these two functions might be very different, as Rob Noble observes in some of his experiments, in that case we will have made more than a measurement. We will have discovered that the experimental fitness function partitions into a direct interaction part — that could come from direct predation, very short-term or short-range media-based signals, spatial competition, or many other microdynamical effects — and a medium-mediated part: 
Finally, does this mean that just by playing with definitions and writing down the replicator equation I have shown that any replating experiment follows replicator dynamics? That would be ridiculous. But we aren’t far. To actually establish this, we need only one more piece: we would need to show that the the replatings have the Markov property. In other words, that if the protocol is kept fixed then from one plate to another then the proportion of type A cells (and maybe a little noise) is all we need to specify the proportion of type A cells that we will have at the end of the grow phase; there is no dependence of plates that are further back in time. Of course, there are many ways this Markov property might break down — and Rob Noble is looking at a recent experiment in a new system where the preliminary data hints towards such a break down.
The easiest way to break the Markov property is if the state-space we consider is insufficient. For example, we assume that the ‘pure’ cultures we start with all have the same strategy. But these cultures were made in a context independent of EGT and the people that purified them probably had different goals than ours in mind. What if the culture is pure for some trait that was of interest to the original experimentalists but heterogeneous in terms of their game behavior. In that case, we need to treat the game as not between type A and type B but between types A, A’ and B where we don’t have an experimental procedure to differentiate between type A and A’. Alternatively, the extra variable we need to add to the state space might not be an extra cell (sub)type but something like a toxin that survives between replatings. This can be done by binding to the surface of the cells — although experimentalists often ‘wash’ cells to avoid this specific effect — or by being absorbed without harm by one cell type to only be actively or passively pumped out of them later and affect the other cell type — a typical example in oncology might be a chemo-resistant cell pumping out the drug after replating and poisoning it’s non-chemo resistant partners. However, all is not lost in this case: there is a rich literature on partially observable Markov decision processes that can come to our rescue (see also footnote [12]).
Of course, expanding the state space might be futile in some cases. The classic examples of this would be cellular or epigenetic memory, certain kinds of phenotype switching, or rapid mutations. In that case, our state space will expand with each replating and it is better to abandon replicator dynamics and adopt some technique that was built for handling historicity. To find this out, though, we would need to first design careful experimental protocols and run our cells and see the long-term behavior breaking the Markov property. Alternatively, if we are lucky and nature cooperates by granting our protocol the Markov property then we will find ourselves in the fortunate position of having a relatively simple operationalist theory of evolutionary games.
Notes and References
- Thank you to reader vzn for bringing it to our attention early. Although I have been looking closely at Archetti’s (2013,2014) theoretical work on non-linear public goods, and even working toward extending it, I would not have noticed this new paper until Google Scholar or a colleague notified me a couple of days after vzn’s comment.
- Note that the exact content of the serum in medium is usually not fully understood for something like the fetal bovine serum that the authors use. In particular, it probably contains chemicals like the IGF-II that is central to the study, but the authors are aware of this and this is why they are so self-conscious about their use of the medium.
- This step is not perfectly characterized, since we might be removing more than just the cells, but we’ll use the auxiliary hypothesis that only (and all) cells are filtered out, all relevant macro-molecules are allowed to pass through, and no contaminant is introduced.
- Producers (abbreviated as +/+ in the paper) come from a beta-tumour cell derived from unsulinomas in Rip1Tag2 mice (Hanahan, 1985), and nonproducers (abbreviated as -/-) come from the same mice with a homozygous deletion of the IGF-II gene (Christofori et al., 1994). Note that here we have an auxiliary hypothesis that the only relevant effect of knocking out the IGF-II gene is that production of IGF-II stops — this is taking a very linear perspective of gene interaction networks, but not something that I want to nit-pick on beyond this mention.
- Rob Noble tells me that his collaborators are running experiments like the one I am describing here, so hopefully those results will be available soon.
- Rob Noble explained to me that although flow cytometry is a standard procedure, it is still one that has important fitness considerations. In particular the plasmid-introduced florescent genes divert some of the energy of the cell to making florescent molecules and thus tend to reduce their fitness relative to that of their non-florescent ancestors. It is also possible to use the florescence to make more detailed measurements than just waiting until replating. For example, Rob’s collaborators film the cells as they are growing, giving hour-by-hour estimates of the (sub)population size from the level of florescence. I will not discuss this sort of experiment in this article, partially because I don’t have too much familiarity with it and partially because it urges people too much toward reductionist stories and thus makes taking a purely operationalist stance more difficult. An example of where more data can make life more difficult rather than less.
- Unfortunately, this is one of the places where the microdynamical replicator dynamics is known to be unreliable. It is tempting to use this to conclude, as Archetti really wants, that we thus have a non-linear public good. But, if we look at ecological games — where both the proportions and the exact numbers of the populations matter — then it is possible to have a heterogeneous equilibrium even with a linear public good (Hauert et al., 2006, 2008). For a more experimental discussion, see Sanchez & Gore (2013; also the introduction by Allen & Nowak, 2013) that we read for EGT Reading Group 45. Of course, if one opens the spatial structure can-of-worms — as Archetti et al. (2015) do at the end of their paper — then almost anything goes and you can have a multitude of different ways to have heterogeneous equilibria with linear public goods.
- Here it is tempting to say that I am contradicting much of my own philosophy. In particular, I often advocate for qualitative classifications and predictions over numerical accuracy in evolutionary game theory. However, it is important to note that this is something I advocate for heuristic models — which just happen to be most of the EGT models I and others work with — but once you start designing experiments and models together you are no longer working with heuristics, you are in the world of abductions or insilications. In this case, it becomes important to start tightly coupling your theory and experiment and I am perfectly happy to make the transition from heuristics to abductions. But while we are doing so, we must remain mindful that the fit is specific to our experimental protocol and not necessarily other experiments where people have used similar names for their variables.
- Here I wish that the supplemental materials — usually a gold mine of results compared to the main paper for high impact journals — would include more discussion of the experiments, instead of focusing on the mathematical modeling. As far as I can tell, at least 3 different growing times were used for the 3 figures, making them incommensurable. The ‘Proliferation Assay’ of the ‘Materials and Methods’ section talks about a relative change in density after a 10 day growth, so I assume that is the growth time for figure 1. On the other hand, figure 2 has data points about every 7 days, and figure 3 talks about 5 days. The description of the proliferation assay suggests that 10 days keeps the cells in the log phase so we can assume that all 3 lengths of growth can be translated between by calculating the slope from them on a log-plot and using that as the comparison rate. However, this forces us to introduce an extra auxiliary hypothesis — that the sigmoidal growth curve of the cells didn’t reach it’s ‘stationary phase’ and that such phases can be cleanly separated — which could have been avoided by just using constant experimental procedures between the figures allowing the word ‘growth rate’ or ‘fitness’ to have the same operational meaning in all 3 figures.
- For a typical replating procedure, we would use a fixed mix of serum for every new plate. However, if we want to compared to the mixed media experiments — as I will want to further down — then we will have the mix depend on the proportion p of type A cells at the end of the previous plate — that is why it is important to do flow cytometry at every replating. In particular, the new serum will be a mix of p parts of media conditioned by type A cells to 1 – p parts of media conditioned by type B cells.
- In some ways, this can be seen as the exorcism of metaphor from science, something that I would not generally endorse since I believe metaphor is central to any kind of understanding. The exception is times of great ontological confusion, where we have so many things that might be relevant and we have no real way — beyond an often faulty and systematically biased intuition — to figure out which to focus on. Once operationalism gives us some ground to stand on, we can go back to reductionism and metaphors as usual. For more, see footnote [8].
- In this equation, some readers might also see Bayes’ rule or just a general occurrence of the multiplicative weight update algorithm that props up eerily often in evolution and learning (Arora et al., 2012; Barton et al., 2014). We should not dismiss this as coincidence, since this observation combined with a consistent used of the logic transform might allow us to exploit the rich theory of learning Markov Decision Processes to help us fit to data in more difficult cases than two-strategy replicator dynamics.
- These last two sentences — and how they allow us to hide the pedantics of microdynamics inside of how we measure — is the crux of my response to Philip Gerlee and Philipp Altrock. Unfortunately, for this post I will leave it at this cryptic comment that hopefully Philip and Philipp can parse, but I promise that in the future I will expand this idea into a more carefully justified post.
- Although even here, we might want to allow for some low complexity interpolation between the two functions, like proportionality, log-proportionality, or logit-proportionality and accept the approximate equality if the two functions are equal under this comparison instead of strict numerical equality which would correspond to the simplest map: identity.
-
To really convince me that we have IGF-II as a public good, the authors would need to redo the mixed media experiments, but instead of combining a proportion p of producer conditioned medium to a proportion 1 – p of nonproducers conditioned medium, they would need need to add to nonproducer conditioned medium variable amounts of IGF-II according to some function f(p) to produce the fitnesses
. If we have
for some (reasonable) monotonically increasing f then I would be convinced. Notice that Archetti et al. (2015) clearly have the experimental control to do this, since in figure S1 of the supplemental material they produce the graph for
. Strangely, they don’t make a similar graph for producers, making me suspicious that it doesn’t make a pretty sigmoid. Also, note that this would convince me that IGF-II is a public good, not that an operationalized version of Archetti’s nonlinear public goods game microdynamics are a good model of this public good. That would require the graphs to not only agree but also having the right functional form.
-
Again, the best way to partition might be something other than ‘+’ (that corresponds to identity or proportionality) and we might use another low complexity function to partition like log (
) or logit (
). Whatever choice is made, should be consistent with the choice for approximation in footnote [14] so that the domain to which the different Fs map have a consistent mathematical structure.
Allen, B., & Nowak, M. A. (2013). Cooperation and the fate of microbial societies. PLoS Biology, 11(4): e1001549.
Archetti, M. (2013). Evolutionary game theory of growth factor production: implications for tumour heterogeneity and resistance to therapies. British Journal of Cancer, 109(4): 1056-1062.
Archetti, M. (2014). Evolutionary dynamics of the Warburg effect: glycolysis as a collective action problem among cancer cells. Journal of Theoretical Biology, 341: 1-8.
Archetti, M., Ferraro, D.A., & Christofori, G. (2015). Heterogeneity for IGF-II production maintained by public goods dynamics in neuroendocrine pancreatic cancer. Proceedings of the National Academy of Sciences of the United States of America, 112 (6), 1833-8 PMID: 25624490
Arora, S., Hazan, E., & Kale, S. (2012). The Multiplicative Weights Update Method: a Meta-Algorithm and Applications. Theory of Computing, 8(1): 121-164.
Barton, N. H., Novak, S., & Paixão, T. (2014). Diverse forms of selection in evolution and computer science. Proceedings of the National Academy of Sciences, 111(29): 10398-10399.
Crespi, B. J. (2001). The evolution of social behavior in microorganisms. Trends in Ecology & Evolution, 16(4): 178-183.
Christofori, G., Naik, P., & Hanahan, D. (1994). A second signal supplied by insulin-like growth factor II in oncogene-induced tumorigenesis. Nature, 369(6479): 414-418.
Gore, J., Youk, H., & Van Oudenaarden, A. (2009). Snowdrift game dynamics and facultative cheating in yeast. Nature, 459(7244): 253-256.
Hanahan, D. (1985). Heritable formation of pancreatic beta-cell tumours in transgenic mice expressing recombinant insulin/simian virus 40 oncogenes. Nature, 315(6015): 115-122.
Hauert, C., Holmes, M., & Doebeli, M. (2006). Evolutionary games and population dynamics: maintenance of cooperation in public goods games. Proceedings of the Royal Society B: Biological Sciences, 273(1600): 2565-2571.
Hauert, C., Wakano, J. Y., & Doebeli, M. (2008). Ecological public goods games: cooperation and bifurcation. Theoretical Population Biology, 73(2): 257.
Sanchez, A., & Gore, J. (2013). Feedback between population and evolutionary dynamics determines the fate of social microbial populations. PLoS Biology, 11(4): e1001547.
Strassmann, J., & Queller, D. (2011). Evolution of cooperation and control of cheating in a social microbe. Proceedings of the National Academy of Sciences, 108(2): 10855-10862
Lenski, R. E., Rose, M. R., Simpson, S. C., & Tadler, S. C. (1991). Long-term experimental evolution in Escherichia coli. I. Adaptation and divergence during 2,000 generations. American Naturalist, 1315-1341.
Lenski, R. E., & Velicer, G. J. (2001). Games microbes play. Selection, 1(1): 89-96.
Ribeck, N., & Lenski, R. E. (2014). Modeling and quantifying frequency-dependent fitness in microbial populations with cross-feeding interactions. bioRxiv, 012807.
Velicer, G. J. (2003). Social strife in the microbial world. Trends in microbiology, 11(7), 330-337.
West, S. A., Diggle, S. P., Buckling, A., Gardner, A., & Griffin, A. S. (2007). The social lives of microbes. Annual Review of Ecology, Evolution, and Systematics, 53-77.
Wiser, M. J., Ribeck, N., & Lenski, R. E. (2013). Long-term dynamics of adaptation in asexual populations. Science, 342(6164): 1364-1367.


Pingback: An update | Theory, Evolution, and Games Group
Pingback: EGT Reading Group 46-50 and a photo | Theory, Evolution, and Games Group
Pingback: Operationalizing the local environment for replicator dynamics | Theory, Evolution, and Games Group
Pingback: Cancer, bad luck, and a pair of paradoxes | Theory, Evolution, and Games Group
Pingback: Abusing numbers and the importance of type checking | Theory, Evolution, and Games Group
Since mine and Philipp’s initial critique of the Archetti-paper I’ve now returned to it with a more constructive mind set.
With regards to this blog post I’m not sure about the relevance of replating for the data in the Archettii-paper. My interpretation of fig. 3C is that no replating was carried out. They simply compare the initial and end fractions of producers. In the Methods it is said that 30 000 cells were plated (in a 24-well plate with growth area 2cm^2 [1]). If we assume that one cell has radius 10 microns then they cover 5 % of the well. Further, if we assume purely exponential growth with an average doubling time of one day we have exceeded the carrying capacity of the well after 5 days. This suggests that competition for space is an important mechanism at work.
The same interpretation applies to fig. 2.
Currently I’m working on a spatially explicit model, but I don’t have any real results to show yet. In any case I think the fact that the population is growing (and competing for space) is of importance if we want to understand the data in the Archetti-paper and also tumour growth, where replating certainly does not occur.
[1] The growth area was found on the manufacturer’s webpage.
Pingback: From linear to nonlinear payoffs in the double public goods game | Theory, Evolution, and Games Group
Pingback: Cataloging a year of blogging | Theory, Evolution, and Games Group
Pingback: Measuring games in the Petri dish | Theory, Evolution, and Games Group
Pingback: Multiple realizability of replicator dynamics | Theory, Evolution, and Games Group
Pingback: Chemical games and the origin of life from prebiotic RNA | Theory, Evolution, and Games Group
Pingback: Cataloging a year of cancer blogging: double goods, measuring games & resistance | Theory, Evolution, and Games Group
Pingback: Ontology of player & evolutionary game in reductive vs effective theory | Theory, Evolution, and Games Group
Pingback: Replicator dynamics and the simplex as a vector space | Theory, Evolution, and Games Group
Pingback: Token vs type fitness and abstraction in evolutionary biology | Theory, Evolution, and Games Group
Pingback: Abstract is not the opposite of empirical: case of the game assay | Theory, Evolution, and Games Group
Pingback: Personal case study on the usefulness of philosophy to biology | Theory, Evolution, and Games Group
Pingback: Danger of motivatiogenesis in interdisciplinary work | Theory, Evolution, and Games Group