This week, I am at the University of Nottingham for the joint meeting of the Society of Mathematical Biology and the European Conference on Mathematical and Theoretical Biology — ECMTB/SMB 2016. It is a huge meeting, with over 800 delegates in attendance, 308 half-hour mini-symposium talks, 264 twenty-minute contributed talks, 190 posters, 7 prize talks, 7 plenary talks, and 1 public lecture. With seventeen to eighteen sessions running in parallel, it is impossible to see more than a tiny fraction of the content. And impossible for me to give you a comprehensive account of the event. However, I did want to share some moments from this week. If you are at ECMTB and want to share some of your highlights for TheEGG then let me know, and we can have you guest post.
I did not come to Nottingham alone. Above is a photo of current/recent Moffitteers that made their way to the meeting this year.
On the train ride to Nottingham, I needed to hear some success stories of mathematical biology. One of the ones that Dan Nichol volunteered was the SIR-model for controlling the spread of infectious disease. This is a simple system of ODEs with three compartments corresponding to the infection status of individuals in the population: susceptible (S), infectious (I), recovered (R). It is given by the following equations
where and are usually taken to be constants dependent on the pathogen, and the total number of individuals is an invariant of the dynamics.
As the replicator dynamics are to evolutionary game theory, the SIR-model is to epidemiology. And it was where Julia Gog opened the conference with her plenary on the challenges of modeling infectious disease. In this post, I will briefly touch on her extensions of the SIR-model and how she used it to look at the 2009 swine flu outbreak in the US. Read more of this post
Last time in the Petri dish, I considered the replicator dynamics between type-A and type-B cells abstractly. In the comments, Arne Traulsen pointed me to Li et al. (2015):
We have attempted something similar in spirit with bacteria. Looking at frequencies alone, it looked like coordination. But taking into account growth led to different conclusions […] In that case, things were more subtle than anticipated…
So following their spirit, I will get more concrete in this post and replace type-A by Curvibacter sp. AEP13 and type-B by Duganella sp. C1.2 — two bacteria that help fresh water Hydra avoid fungal infection. And I will also show how to extend our replicator dynamics with growth and changing cell density.
Although I try to follow Arne’s work very closely, I had not read Li et al. (2015) before, so I scheduled it for a reading group this past Friday. I really enjoyed the experiments that they conducted, but I don’t agree with their interpretations that taking growth into account leads to a different conclusion. In this post, I will sketch how they measured their experimental system and then provide a replicator equation representation of the Lotka-Volterra model they use to interpret their results. From this, we’ll be able to conclude that C and D are playing the Stag Hunt — or coordination, or assurance, pick your favorite terminology — game.
One of my favorite parts of mathematical modeling is the opportunities it provides to carefully explore metaphors and analogies between disciplines. The connection most carefully explored at the MBI Workshop on the Ecology and Evolution of Cancer was, as you can guess from the name, between ecology and oncology. Looking at cancer from the perspective of evolutionary ecology can offer us several insights that the standard hallmarks of cancer approach (Hanahan & Weingerg, 2000) hides. I will start with some definitions for this view, discuss ecological concepts like mimicry and ecological engineers in the context of cancer, unify these concepts through the idea of morphostatic maintenance of tissue microarchitecture, and finish with the practical importance of diet to cancer. Read more of this post
If I had to identify one then my main allergy would be bloated models. Although I am happy to play with complicated insilications, if we are looking at heuristics where the exact physical basis of the model is not established then I prefer to build the simpleast possible model that is capable of producing the sort of results we need. In particular, I am often skeptical of agent based models because they are simple to build, but it is also deceptively easy to have the results depend on an arbitrary data-independent modeling decision — the curse of computing. Therefore, as soon as I saw the agent-based models for the effect of imatinib on stem-cells in chronic myeloid leukemia (Roeder et al., 2002; 2006; Horn et al., 2013 — the basic model is pictured above), I was overcome with the urge to replace it by a simpler system of differential equations. Read more of this post
As you might have guessed from my recent posts, I am cautious in trying to use mathematics to build insilications for predicting, profiting from, or controlling financial markets. However, I realize the wealth of data available on financial networks and interactions (compared to similar resources in ecology, for example) and the myriad of interesting questions about both economics and humans (and their institutions) more generally that understanding finance can answer. As such, I am more than happy to look at heuristics and other toy models in order to learn about financial systems. I am particularly interested in understanding the interplay between individual versus systemic risk because of analogies to social dilemmas in evolutionary game theory (and the related discussions of individual vs. inclusive vs. group fitness) and recently developed connections with modeling in ecology.
Three-month Libor-overnight Interest Swap based on data from Bloomberg and figure 1 of Domanski & Turner (2011). The vertical line marks 15 September 2008 — the day Lehman Brothers filed for bankruptcy.
A particular interesting phenomenon to understand is the sudden liquidity freeze during the recent financial crisis — interbank lending beyond very short maturities virtually disappeared, three-month Libor (a key benchmarks for interest rates on interbank loans) skyrocketed, and the world banking system ground to a halt. The proximate cause for this phase transition was the bankruptcy of Lehman Brothers — the fourth largest investment bank in the US — at 1:45 am on 15 September 2008, but the real culprit lay in build up of unchecked systemic risk (Ivashina & Scharfstein, 2010; Domanski & Turner, 2011; Gorton & Metrick, 2012). Since I am no economist, banker, or trader, the connections and simple mathematical models that Robert May has been advocating (e.g. May, Levin, & Sugihara (2008)) serve as my window into this foreign land. The idea of a good heuristic model is to cut away all non-essential features and try to capture the essence of the complicated phenomena needed for our insight. In this case, we need to keep around an idealized version of banks, their loan network, some external assets with which to trigger an initial failure, and a way to represent confidence. The question then becomes: under what conditions is the initial failure contained to one or a few banks, and when does it paralyze or — without intervention — destroy the whole financial system? Read more of this post
Theoretical physicists have the reputation of an invasive species — penetrating into other fields and forcing their methods. Usually these efforts simply irritate the local researchers, building a general ambivalence towards field-hopping physicists. With my undergraduate training primarily in computer science and physics, I’ve experienced this skepticism first hand. During my time in Waterloo, I tried to supplement my quantum computing work by engaging with ecologists. My advances were met with a very dismissive response:
But at the risk of sounding curmudgeonly, it is my experience that many folks working in physics and comp sci are more or less uninformed regarding the theoretical ecology, and tend to reinvent the wheel.
On rare occasion though, a theorist will move into a field of sledges & rollers, and help introduce the first wheel. This was the case 40 years before my ill-fated courtship of Waterloo ecologists, when Robert May published “Stability in multispecies community models” (1971) and transitioned from theoretical physics (PhD 1959, University of Sydney) to ecology. He helped transform the field from shunning equations to a vibrant community of observation, experiments, and mathematical models.
Lord Robert May of Oxford. Photo is from the donor’s page of Sydney High School Old Boys Union where he attended secondary school.
Robert M. May, Lord May of Oxford, is a professor in the Department of Zoology at University of Oxford. I usually associate him with two accomplishments inspired by (but independent of) ecology. First, he explored the logistic map and its chaotic behavior (May, 1976), becoming one of the co-founders of modern chaos theory. Although the origins of chaos theory can be traced back to another great cross-disciplinary scholar — Henri Poincaré; it wasn’t until the efforts of May and colleagues in the 1970s that the field gained significant traction outside of mathematics and gripped the popular psyche. Second, he worked with his post-doc Martin A. Nowak to popularize the spatial Prisoner’s Dilemma and computer simulation as an approach to the evolution of cooperation (Nowak & May, 1992). This launched the sub-field that I find myself most comfortable in and stressed the importance of spatial structure in EGT. May is pivoting yet again, he is harnessing his knowledge of ecology and epidemiology to study the financial ecosystem (May, Levin, & Sugihara, 2008).
After the 2008 crises, finance became a hot topic for academics and May, Levin, & Sugihara (2008) suggested mathematical ecology as a source of inspiration. Questions of systemic risk, or failure of the whole banking system (as opposed to a single constituent bank), grabbed researchers’ attention. In many ways, these questions were analogous to the failure of ecosystems. In fisheries research there was a similar history to that of finance. Early research on fisheries would fixate on single species, the equivalent of a bank worrying only about its own risk-management strategy. However, the fishes were intertwined in an ecological network like banks are connected through an inter-bank loan network. The external stresses fish species experiences were not independent, something like a change in local currents or temperature would effect many species at once. Analogously, the devaluation of an external asset class like the housing market effects many banks at once. As over-consumption depleted fisheries in spire of ecologists’ predictions, the researchers realized that they must switch to a holistic view; they switched their attention to the whole ecological network and examined how the structure of species’ interactions could aid or hamper the survival of the ecosystem. Regulators have to view systemic risk in financial systems through the same lens by considering a holistic approach to managing risk.
Once a shock is underway, ideas from epidemiology can help to contain it. As one individual becomes sick, he has the risk of passing on that illness to his social contacts. In finance, if a bank fails then the loans it defaulted on can cause its lenders to fail and propagate through the inter-bank loan network. Unlike engineered networks like electrical grids, an epidemiologist does not have control over how humans interact with each other, she can’t design our social network. Instead, she has to deter the spread of disease through selective immunization or through encouraging behavior that individuals in the population might or might not adopt. Similarly, central bankers cannot simply tell all other banks who to loan to, instead they must target specific banks for intervention (say through bail-out) or by implementing policies that individual banks might or might not follow (depending on how these align with their interests). The financial regulator can view bank failure as a contagion (Gai & Kapadia, 2010) and adapt ideas from public health.
The best part of mathematical models is that the preceding commonalities are not restricted to analogy and metaphor. May and colleagues make these connections precise by building analytic models for toy financial systems and then using their experience and tools from theoretical ecology to solve these models. Further, the cross-fertilization is not one-sided. In exchange for mathematical tools, finance provides ecology with a wealth of data. Studies like the one commissioned by the Federal Reserve Bank of New York (Soramäki et al., 2007) can look at the interaction of 9500 banks with a total of 700000 transfers to reveal the topology of inter-bank payment flows. Ecologists can only dream of such detailed data on which to test their theories. For entertainment and more information, watch Robert May’s hour-long snarky presentation of his work with Arinaminpathy, Haldane, and Kapadia (May & Arinaminpathy 2010; Haldane & May, 2011; Arinaminpathy, Kapadia, & May, 2012) during the 2012 Stanislaw Ulam Memorial Lectures at the Santa Fe Institute:
References
Arinaminpathy, N., Kapadia, S., & May, R. M. (2012). Size and complexity in model financial systems. Proceedings of the National Academy of Sciences, 109(45), 18338-18343.
Gai, P., & Kapadia, S. (2010). Contagion in financial networks. Proceedings of the Royal Society A: Mathematical, Physical and Engineering Science, 466(2120), 2401-2423.
Haldane, A. G., & May, R. M. (2011). Systemic risk in banking ecosystems. Nature, 469(7330), 351-355.
May, R. M. (1971). Stability in multispecies community models. Mathematical Biosciences, 12(1), 59-79.
May, R. M. (1976). Simple mathematical models with very complicated dynamics. Nature, 261(5560), 459-467.
May RM, Levin SA, & Sugihara G (2008). Ecology for bankers. Nature, 451 (7181), 893-5 PMID: 18288170
May, R. M., & Arinaminpathy, N. (2010). Systemic risk: the dynamics of model banking systems. Journal of the Royal Society Interface, 7(46), 823-838.
Nowak, M. A., & May, R. M. (1992). Evolutionary games and spatial chaos. Nature, 359(6398), 826-829.
Soramäki, K., Bech, M. L., Arnold, J., Glass, R. J., & Beyeler, W. E. (2007). The topology of interbank payment flows. Physica A: Statistical Mechanics and its Applications, 379(1), 317-333.

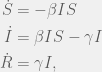





 and its chaotic behavior (May, 1976), becoming one of the co-founders of
and its chaotic behavior (May, 1976), becoming one of the co-founders of 

Ecology of cancer: mimicry, eco-engineers, morphostats, and nutrition
October 9, 2014 by Artem Kaznatcheev 8 Comments
One of my favorite parts of mathematical modeling is the opportunities it provides to carefully explore metaphors and analogies between disciplines. The connection most carefully explored at the MBI Workshop on the Ecology and Evolution of Cancer was, as you can guess from the name, between ecology and oncology. Looking at cancer from the perspective of evolutionary ecology can offer us several insights that the standard hallmarks of cancer approach (Hanahan & Weingerg, 2000) hides. I will start with some definitions for this view, discuss ecological concepts like mimicry and ecological engineers in the context of cancer, unify these concepts through the idea of morphostatic maintenance of tissue microarchitecture, and finish with the practical importance of diet to cancer.
Read more of this post
Filed under Commentary, Reviews Tagged with Alan Turing, conference, ecology, mathematical oncology