Everyday I walk to the Stabile Research Building to drink espresso and sit in my cozy — although oversaturated with screens — office. Oh, and to chat about research with great people like Arturo Araujo, David Basanta, Jill Gallaher, Jacob Scott, Robert Vander Velde and other Moffitters. This walk to the office takes about 30 minutes each way, so I spend it listening to podcasts. For the past few weeks, upon recommendation from a friend, I’ve started listing to the archive of the Very Bad Wizards. This is a casual — although oversaturated with rude jokes — conversation between David Pizarro and Tamler Sommers on various aspects of the psychology and philosophy of morality. They aim at an atmosphere of two researchers chatting at the bar; although their conversation is over Skype and drinks. It is similar to the atmosphere that I want to promote here at TheEGG. Except they are funny.
To do the quiz, you are asked to rate 12 statements (well, 11 and one question about browsers) on a six point Likert scale from strongly disagree to strongly agree. Here are the three that caught my attention:
If I heard that a new restaurant in my neighborhood blended the cuisines of two very different cultures, that would make me want to try it.
My government should treat lives of its citizens as being much more valuable than lives in other countries.[3]
I wish the world did not have nations or borders and we were all part of one big group.[4]
Do you strongly agree? Strongly disagree? What was your overall place on the liberal-conservative scale?
Regardless of your answers, the statements probably remind you of an important aspect of your daily experience. The world is divided into a diversity of groups, and they coexist in a tension between their arbitrary, often artificial, nature and the important meaning that they hold to both their own members and others. Often this division is accompanied by ethnocentrism — a favoring of the in-group at the expensive of, or sometimes with direct hostility toward, the out-group — that seems difficult to circumvent through simply expanding our moral in-group. These statements also confront you with the image of what a world without group lines might look like; would it be more cooperative or would it succumb to the egalitarian dilemma?[5]
As you know, dear reader, here at TheEGG we’ve grappled with some of these questions. Mostly by playing with the Hammond & Axelrod model of ethnocentrism (2006; also see: Hartshorn, Kaznatcheev & Shultz, 2012). Recently, Jansson’s (2015) extension of my early work on the robustness of ethnocentrism (Kaznatcheev, 2010) has motivated me to continue this thread. A couple of weeks ago I sketched how to reduce the dimensionality of the replicator equations governing tag-based games. Today, I will use this representation to look at how properties of the game affect the persistence and diversity of tags. Read more of this post
Mathematicians and physicists love finding symmetries. The reason is simple: symmetries make life easier. The situation is no different when studying the evolutionary dynamics of life. If the fitness functions of your organisms have some symmetry or other nice structure then you can usually exploit it to make analyzing your replicator equations easier. In this post, I want to show an example of this in tag-based models. This analysis is an essential base case when building more complicated models of ethnocentrism — like our work in the Hammond and Axelrod model — and I have been meaning to write about it for a while. This will give me a chance to show a concrete example where my method for factoring the replicator equation is useful, and how observing a straighforward symmetry can reduce the dimensionality of a problem. Maybe this exercise will also teach us something about the evolution of ethnocentrism. Read more of this post
Yesterday was the first day of the 4th Integrated Mathematical Oncology Workshop here at Moffitt. This year, it is run jointly with the Center for Infection Research in Cancer and is thus focused on the interaction of infection disease and cancer. This is a topic that I have not focused much attention on — except for the post on canine transmissible venereal tumor and passing mentions of Human papillomavirus (HPV) — so I am excited for the opportunity to learn. The workshop opened with a half-day focused on getting to know the external visitors, Alexander Anderson’s introduction, and our team assignments. I will be teammates with Heiko Enderling, Domenico Coppola, Jose M. Pimiento, and others. We will be looking at Helicobacter pylori. Go team blue! If you are curious, the more popularly known HPV went to David Basanta’s team, it will be great to compete against my team leader from last year. As you can expect, the friendly trash talking and subtle intimidation has already begun.
To be frank, before yesterday, I’ve only ever heard of H. pylori once and knew nothing of its links to stomach cancer. The story I heard was associated with Barry J. Marshall and J. Robin Warren’s award of the 2005 Nobel Prize in Physiology and Medicine “for their discovery of the bacterium Helicobacter pylori and its role in gastritis and peptic ulcer disease”. In 1984, Marshall was confident in the connection between H. pylori, inflammation, and ulcers, but the common knowledge of the day was that ulcers were caused by things like stress and smoking, not bacteria. The drug companies even happened to have an expensive drug that could manage the associated stomach inflammation, and given the money it was bringing in, nobody was concerned with finding some bacterium that could be cured with cheap antibiotics. Having difficulty convincing his colleagues (apart from Warren), Marshall decided to drink a Petri dish of cultured H. pylori, and within a few days grew sick, developing severe inflammation of the stomach before finally (two weeks after the ingestion) going on antibiotics and curing himself. This dramatic display was sufficient to push for bigger studies that eventually lead to the Nobel prize; I recommend listening to Warren’s podcast with Nobel Prize Talks or his acceptance speech for the whole story.
This is a fascinating tale, but from the modeling perspective, the real excitement of H. pylori and its role in stomach cancer is the multitude of scales that are central to the development of disease. We see important players from the scale of molecules involved in changing stomach acidity, to the single-cell scale of the bacteria and stomach lining, to the changes across the stomach as a whole organ, and the role of the individual patient’s life style and nutrition. These are the usual scales we see when modeling cancer, and dovetail nicely with Anderson’s opening remarks on the centrality of mathematics in helping us bridge the gaps. However, in the case of H. pylori, the scales go beyond the single individual at which Anderson stops and extend to the level of populations of humans in the co-evolution of host and pathogen, and even populations of groups of humans in a speculative connection to a topic familiar to TheEGG readers — the evolution of ethnocentrism. In preparation for the second half of the second day and the intense task of finding a specific question for team blue to focus on, I wanted to give a quick overview of these scales. Read more of this post
Running an agent based simulation really isn’t that complex. While there’s no shortage of ready-made software packages for ABM (like Repast and NetLogo), all you really need is a good, high-level programming language and a code editor.
As you may have noticed from other blog posts, we have spent quite a bit of time studying agent based models of ethnocentric evolution. To coincide with the publication of our paper (Hartshorn, Kaznatcheev & Shultz, 2013) on the evolution of ethnocentrism in the Journal of Artificial Societies and Social Simulation (JASSS), we thought it would be fun to provide a hands-on tutorial so you can replicate the model yourself. There’s a lot to cover here, so we won’t get into the scientific description of the model itself, but you can read a good synopsis in my executive summary, or Artem’s general overview.
This post assumes no programming background, just a computer, patience, and some curiosity. That being said, you will be compiling a small Java program and modifying its source code, so if words like “compile,” “source code,” and “Java” strike terror in your heart, consider yourself forewarned. It’s actually not that scary. In Estonia they’re teaching kids to program in first grade, and you’re smarter than a first grader…right?! Read more of this post
Cooperation is a paradox—it just doesn’t make sense. Why should I help you when there’s no direct benefit for me? Artem, Professor Tom Shultz, and I have been working for quite some time on a paper about cooperation, and we’re psyched to announce that it’s finally been published in The Journal of Artificial Societies and Social Simulation (JASSS). JASSS is an open web journal, so you can view the full text of our article for free on their website. Or you could skip the 8000 or so words and check out this summary post. Read more of this post
I haven’t been delving into evolutionary game theory and agent-based modeling for very long, and yet I find that in that little time something quite eerie happens once I’m immersed in these models and simulations: I find myself oscillating between two diametrically opposed points of view. As I watch all of these little agents play their games using some all-too-simplistic strategy, I feel like a small God*. I watch cooperators cooperate, and defectors defect oblivious to what’s in their best interest at the moment. Of course, in the end, my heart goes out to the cooperators, who unfortunately can’t understand that they are being exploited by the defectors. That is what pushes me at the other end of the spectrum of omniscience, and with a nudge of empathy I find myself trying to be a simpleton agent in my over-simplified world.
In that state of mind, I begin to wonder what information exists in the environment, in particular information about the agents I am going to play against. I suppose I’m able to access it and use it to condition my move. Admittedly, that makes me a bit more complex than my original simpleton, and that complexity is likely to come at a cost, but I leave it to evolution to figure out whether the trade-off is worthwhile. Read more of this post
Artem Kaznatcheev and I presented a poster on May 4th at the University of British Columbia to a highly interdisciplinary conference on religion. The conference acronym is CERC, which translates as Cultural Evolution of Religion Research Consortium. Most of the 60-some attendees are religion scholars and social scientists from North American and European universities. Many are also participants in a large partnership grant from the Social Sciences and Humanities Research Council of Canada (SSHRC), spearheaded by Ted Slingerland, an East Asian scholar at UBC. Some preliminary conversations with attendees indicated considerable apprehension about how researchers from the humanities and sciences would get on. Many of us are familiar with collaborative difficulties even in our own narrow domains. Skepticism was fairly common.
As far as I know, our poster was the only computer simulation presented at the meeting. We titled it Agent-based modeling of the evolution of “religion”, with scare quotes around religion because of the superficial and off-hand way we treated it. Because we know from experience that simulations can be a tough sell even at a scientific psychology conference, we were curious about whether and how this poster would fly in this broader meeting. Read more of this post
We have previously discussed the importance of population structure in evolutionary game theory, and looked at the Ohtsuki-Nowak transform for analytic studies of games on one of the simplest structures — random regular graphs. However, there is another extremely simple structure to consider: a family of inviscid sets. We can think of each agent as belonging to one or more sets and interacting with everybody that shares a set with them. If there is only one set then we are back to the case on a completely inviscid population. If we associate a set with each edge in a graph and restrict them to have constant size then we have standard evolutionary graph theory. However, it is more natural to allow sets to grow larger if their members have high fitness.
Tarnita et al. (2009) consider a population of individuals and sets, where each individual can belong to of the sets. Strategy and set membership are heritable (with mutation probabilities and , respectively), and interactions are only with agents that share a set (if two agents share more than one set then they interact more than once). However, reproduction is inviscid: a random individual is selected to die and everybody competes to replace them with a child. This set-dependent interaction, makes the model equivalent to the earliest models of ethnocentrism, but the model is not equivalent to more modern approaches to ethnocentrism. Since sets cannot reproduce and migration (through mutation) between sets is purely random, the model also cannot capture group selection. However, cooperation for the Prisoners’ dilemma still emerges in this model, if we have:
Where is the population-scaled set mutation rate, even when this is zero we get cooperation when . Alternatively, to simplify we can take the limit of to get:
If we allow the maximum number of sets () and take the further limit of large populations then this becomes a very simple:
Finally, in the supplementary materials, the authors derive a very nice relationship for evolution of cooperation on arbitrary cooperate-defect games given by a payoff matrix of:
where is a structural constant given by:
Which in the limit of small population-scaled strategy mutation (), and becomes:
Since the structural constant is always greater than 1, and since we typically care about large , it is more enlightening to look at the reciprocal that in the limit of large becomes:
And simplifies the general equation for cooperation to emerge to:
Which can be seen as a relaxation of the classic game theory concept of risk-dominance.
Tarnita, C., Antal, T., Ohtsuki, H., & Nowak, M. (2009). Evolutionary dynamics in set structured populations Proceedings of the National Academy of Sciences, 106 (21), 8601-8604 DOI: 10.1073/pnas.0903019106
Ethnocentrism (or contingent altruism) can be viewed as one of many mechanisms for enabling cooperation. The agents are augmented with a hereditary tag and the strategy space is extended from just cooperation/defection to behaviour that can be contingent on if the diad share or differ in their tag. The tags and strategy are not inherently correlated, but can develop local correlations due to system dynamics. This can expand the range of environments in which cooperation can be maintained, but an assortment-biasing mechanism is needed to fuel the initial emergence of cooperation (Kaznatcheev & Shultz, 2011). The resulting cooperation is extended only towards the in-group while the out-group continues to be treated with the cold rationality of defection.
Suppose that circles are the in-group and squares the out-group. The four possible strategies and their minimal representations as finite state machines is given.
The four possible strategies are depicted above, from top to bottom: humanitarian, ethnocentric, traitorous, and selfish. Humanitarians and selfish agents do not condition their behavior on the tag of their partner, and do not require the cognitive ability to categorize. Although this ability is simple, it can still merit a rich analysis (see: Beer, 2003) by students of minimal cognition. By associating a small fitness cost with categorization, we can study how much ethnocentric (and traitorous) agents are willing to pay for their greater cognitive abilities. This cost directly changes the default probability to reproduce (), with humanitarians and selfish agents having and ethnocentrics and traitorous agents having . During each cycle, the is further modified by the game interactions, with each cooperative action costing and providing a benefit (that varies depending on the simulation parameters) to the partner. For more detailed presentation of the simulation and default parameter, or just to follow along on your computer, I made my code publicly available on GitHub. Pardon its roughness, the brunt of it is legacy code from when I first build this model in 2009 for Kaznatcheev (2010).
Number of agents by strategy versus evolutionary cycle. The lines represent the number of agents of each strategy: blue — humanitarian; green — ethnocentric; yellow — traitorous; red — selfish. The width of the line corresponds to standard error from averaging 30 independent runs. The two figures correspond to different costs of cognition. The left is k = 0.002 and is typical of runs before the cognitive cost phase transition. The right is k = 0.007 and is typical of runs after the cognitive cost phase transition. Figure is adapted from Kaznatcheev (2010).
The dynamics for low are about the same as the standard no cognitive cost model as can be seen from the left figure above. However, as increases there is a transition to a regime where humanitarians start to dominate the population, as in the right figure above. To study this, I ran simulations with a set ratio and increasing from 0.001 to 0.02 with steps of 0.001. You can run your own with the command bcRun(2.5,0.001*(1:20)); some results are presented below, your results might differ slightly due to the stochastic nature of the simulation.
Proportion of humanitarians (blue), ethnocentrics (red), and cooperative interactions (black) versus cognitive cost for b/c = 2.5. Dots are averages from evolutionary cycles 9000 to 10000 of 10 independent runs. The lines are best-fit sigmoids and the dotted lines mark the steepest point; I take take this as the point for the cognitive cost phase transition. Data generated with bcRun(2.5,0.001*(1:20)) and visualized with bcPlot(2.5,0.001*(1:20),[],1)
Each data-point is the average from the last 1000 cycles of 10 independent simulations. The points suggest a phase transition from a regime of few humanitarians (blue), many ethnocentrics (red), and very high cooperation (black) to one of many humanitarians, few ethnocentrics, and slightly less cooperation. To get a better handle on exactly where the phase transition is, I fit sigmoids to the data using fitSigmoid.m. The best-fit curves are shown as solid lines; I defined the point of phase transition as the steepest (or inflection) point on the curve and plotted them with dashed lines for reference. I am not sure if this is the best approach to quantifying the point of phase transition, since the choice of sigmoid function is arbitrary and based only on the qualitative feel of the function. It might be better to fit a simpler function like a step-function or a more complicated function from which a critical exponent can be estimated. Do you know a better way to identify the phase transition? At the very least, I have to properly measure the error on the averaged data points and propogate it through the fit to get error bounds on the sigmoid parameters and make sure that — within statistical certainty — all 3 curves have their phase transition at the same point.
The most interesting feature of the phase transition, is the effect on cooperation. The world becomes more equitable; agents that treat out-groups differently from in-group (ethnocentrics) are replaced by agents that treat everyone with equal good-will and cooperation (humanitarians). However, the overall proportion of cooperative interactions decreases — it seems that humanitarians are less effective at suppressing selfish agents. This is consistent with the free-rider suppression hypothesis that Shultz et al. (2009) believed to be implausible. The result is egalitarians’ dilemma: by promoting equality among agents the world becomes less cooperative. Should one favour equality and thus individual fairness over the good of the whole population? If we expand our moral circle to eliminate out-groups will that lead to less cooperation?
In the prisoners’ dilemma, we are inclined to favor the social good over the individual. Even though it is rational for the individual to defect (securing a higher payoff for themselves than cooperating), we believe it is better for both parties to cooperate (securing a better social payoff than mutual defection). But in the egalitarians’ dilemma we are inclined to favour the individualistic strategy (fairness for each) over the social good (higher average levels of cooperative interactions). We see a similar effect in the ultimatum game: humans reject unfair offers even though that results in neither player receiving a payoff (worse for both). In some ways, we can think of the egalitarians’ dilemma as the population analogue of the ultimatum game; should humanity favor fairness over higher total cooperation?
I hinted at some of these questions in Kaznatcheev (2010) but I restrained myself to just . From this limited data, I concluded that since the phase transition happens for less than any other parameter in the model, it must be the case that agents are not willing to invest much resources into developing larger brains capable of categorical perception just to benefit from an ethnocentric strategy. Ethnocentrism and categorical perception would not have co-evolved, the basic cognitive abilities would have to be in place by some other means (or incredibly cheap) and then tag-based strategies could emerge.
Value of k at phase transition versus b/c ratio. In blue is the transition in proportion of humanitarians, red — proportion of ethnocentrics, and black – proportion of cooperative interactions. Each data point is made from a parameter estimate done using a sigmoid best fit to 200 independent simulations over 20 values of k at a resolution of 0.001.
Here, I explored the parameter space further, by repeating the above procedure while varying the ratio by changing from 0.02 to 0.035 in increments of 0.0025 while keeping fixed at 0.01. Unsurprisingly, the transitions for proportion of ethnocentrics and humanitarians are indistinguishable, but without a proper analysis it is not clear if the transition from high to low cooperation also always coincides. For , agents are willing to invest more than before the phase transition to all humanitarians, this invalidates my earlier reasoning. Agents are unwilling to invest much resources in larger brains capable of categorical perception only for competitive environments (low ). As increases, the agents are willing to invest more in their perception to avoid giving this large benefit to the out-group. This seems consistent with explicit out-group hostility that Kaznatcheev (2010b) observed in the harmony game. However, apart from simply presenting the data, I can’t make much more sense from this figure. Do you have any interpretations? Can we learn something from the seemingly linear relationship? Does the slope (if we plot versus then it is about 0.5) tell us anything? Would you still conclude that co-evolution of tag-based cooperation and categorical perception is unlikely?
References
Beer, Randall D. (2003). The Dynamics of Active Categorical Perception in an Evolved Model Agent. Adaptive Behavior. 11(4): 209-243.
Kaznatcheev, Artem (2010). The cognitive cost of ethnocentrism Proceedings of the 32nd annual conference of the cognitive science society
Kaznatcheev, A. (2010b). Robustness of ethnocentrism to changes in inter-personal interactions. Complex Adaptive Systems – AAAI Fall Symposium.
Kaznatcheev, A., & Shultz, T. R. (2011). Ethnocentrism maintains cooperation, but keeping one’s children close fuels it. Proceedings of the 33rd Annual Conference of the Cognitive Science Society. 3174-3179.
Shultz, T. R., Hartshorn, M., & Kaznatcheev, A. (2009). Why is ethnocentrism more common than humanitarianism? Proceedings of the 31st Annual Conference of the Cognitive Science Society. 2100-2105.
Any good story starts with a colourful character, a complicated character, and — to be complacent with modern leftist literature — an anarchist intellectual well-versed in (but critical and questioning of) Marxism; enter Pyotr Alexeyevich Kropotkin. Today he is best known as one of the founders and leading theorist of anarcho-communism, but in his time he was better known as an anti-Tsarist revolutionary, zoologist, geographer and explorer. Kropotkin was born to the Prince of Smolensk, a descendant of the Rurik dynasty that ruled and eventually unified many of the Principalities and Duchies of Rus into the Tsardom of Russia. By the 9 December 1842 birth of our protagonist, Russia had been under Romanov rule for over 200 years, but the house of Rurik still held great importance. Even though the young boy renounced his Princely title at age 12, he was well-off and educated in the prestigious Corps of Pages. There he rose to the highest ranks and became the personal page of Tsar Alexander II. Upon graduation this entitled Kropotkin to his choice of post, and our first plot twist.
Analogous to the irresistible pull of critical theory on modern liberal-arts students, the young Kropotkin was seduced by the leftist thought of his day: French encyclopédistes, the rise of Russian liberal-revolutionary literature, and his personal disenfranchisement with and doubt of the Tsar’s “liberal” reputation. Instead of choosing a comfortable position in European Russia, the recent graduate requested to be sent to the newly annexed Siberian provinces and in 1862 was off to Chita. This city has a personal significance to me, it is where my grandfather was stationed over 100 years later and most of my mother’s childhood was spent there. Chita has become a minor place of pilgrimage for modern anarchists, but it (and the other Siberian administrative centre at Irkutsk) did not hold Kropotkin’s attention for long.
Unable to enact substantial change as an administrator, he followed his passion as a naturalist. In 1864, he took command of a geographic survey expedition into Manchuria. Having read Darwin’s On the Origin of Species when it was published 5 years earlier, Kropotkin embarked on a distinctly Siberian variant of the HMS Beagle — sleigh dogs instead of wind to power his way. His hope was to observe the same ‘tooth and claw’ competition as Darwin, but instead he saw primarily cooperation. In the harsh environment of Siberia, it wasn’t a struggle of beast versus beast, but animal against environment.
From 1890 to 1896, Kropotkin published his Siberian observations as a series of essays in the British monthly literary magazine, Nineteenth Century. Motivated as a response to Huxley’s “The Struggle for Existence”, the essays highlighted cooperation among nonhuman animals, in primitive societies and medieval cities, and in contemporary times. He concluded that not competition, but cooperation, were the most important factors in survival and the evolution of species. Kropotkin assembled the essays into book form, and in 1902 published Mutual Aid: A Factor of Evolution. A magnum opus on cooperation, much like E.O. Wilson Sociobiology of nearly 75 years later, Kropotkin started from the social insects and traced a common thread to the human society around him; he was the first student of cooperation.
Unfortunately, his mechanism for cooperation did not extend beyond group selection. Kropotkin left it to modern researchers to find more basic engines of altruism. Only now are we starting to build mathematical, computational, and living models. To study cooperation in the laboratory, especially when looking at the effect of environmental austerity, Strassmann & Queller (2011) have proposed the social microbe Dictyostelium discoideum or slime mold as the perfect model. These single-cell soil-dwelling amoeba are capable of working together under austere conditions, and even display rudimentary swarm intelligence. A long time expert on slime molds, John Bonner of Princeton University, made a video of them during his undergraduate years at Harvard:
Under plentiful conditions, D. discoideum are solitary predators of bacteria, which they consume by engulfment. If the environment deteriorates and the amoebae begin to starve, then they enter a social stage. Using their quorum-sensing mechanism they check if enough other amoebae are present in the area and then aggregate into a mound. They coat themselves with a slime (that gives them their name) and move together as a unit, until they find a good location to fruit. The slime mold then extends a stalk up from the soil with most cells forming a spore at the top. At a certain height, the spore is released, allowing the amoebae at the top to disperse to greener pastures; the cells in the stalk die. Since all the cells are free-living independent organisms during the non-social stage, this shows the clearest form of altruism: fellow D. discoideum sacrificing their own lives in order to give their brethren a chance at a future.
Sadly, most evolutionary game theory models assume constant population size and no resource variability. In these models, it is difficult to introduce a parameter analogous to environmental austerity. To allow for resource limitations, we need to introduce variable population sizes and thus create an ecological game. I explored this modification for a Hammond & Axelrod-like model back in the summer of 2009 and thought I would share some results here.
The agents inhabit a toroidal lattice, and each round the agents interacts with their 4 adjacent neighbours via the Prisoner’s dilemma. The payoffs are added to their default birth rate, reproduction is asexual and into adjacent empty sites. At each time step, each agent has a fixed (0.25) probability of expiring and vacating its site. The worlds start empty and are gradually filled with agents.
This figure has three graphs; in each figure the line thickness represents standard error from averaging 30 independent runs. The leftmost graph is the proportion of cooperation versus cycle, with two conditions for default birth rate: 0.24 (high austerity; top line) and 0.28 (low austerity; bottom line). The two figures on the right show the total number of cooperators (blue) and defectors (red). The rightmost graph has time flowing from right to left. The left panel is high austerity (def ptr = 0.24) and the right panel is low austerity (def ptr = 0.28).
Above are the results for a Prisoner’s dilemma interaction with — a rather competitive environment. Matching Shultz, Hartshorn, & Kaznatcheev (2009) and consistent with Milbiner, Cremer, & Frey (2010), we can see an early spike in the number of cooperators as the world reaches its carrying capacity. After this transient period, the dynamics shift and defection becomes more competitive. The dynamics settle to a stable distribution of cooperators and defectors. The proportion of cooperation depends heavily on the environmental austerity. In a harsh environment with a low default birth rate of 0.24, the agents band together and cooperate and in a plentiful environment with high default birth rate of 0.28, defection dominates. As Kropotkin observed: cooperation is essential to surviving environmental austerity.
Analogous to the results from the Hauert, Homles, & Doebeli (2006) ecological public-goods game the proportion of cooperation tends to bifurcate around default birth rate equal to to the death rate (0.25), although I don’t present the visuals here. The increase in default birth rate results in a slight increase in the world population at saturation, but even by raw number there are more cooperators in the high austerity than the low austerity setting. Thus, it is not simply defectors benefiting more from the decrease in austerity (since defectors go from a regime where clusters are not-self sustaining (def ptr = 0.24) to one where it is (def ptr = 0.28)), but also an effect of defectors out-competing and disproportionately exploiting and crowding out the cooperators.
Each graph is evolutionary cycles versus proportion of cooperation, line thickness is standard error from averaging 30 independent runs. Environmental austerity decreases from the left graph (where default birth rate is equal to death rate) to the right (where their ratio is 1.1). The blue line is the model where agents can discriminate based on arbitrary non-strategy related tag (the green-beard effect/ethnocentrism are possible) and the green line is simulations where no conditional strategy is possible.
If agents are allowed to condition their behavior on an arbitrary tag then the ethnocentric population is better able to maintain higher levels of cooperation as environmental austerity decreases. In the tag-based model, it would be interesting to know if there is a parameter range where varying environmental austerity can take us from a regime of humanitarian (unconditional cooperator) dominance, to ethnocentric dominance (cooperate with in-group, defect from out-group), to a selfish (unconditional defection) world. I am also curious to know how the irrational hostility I observed in the tag-based harmony game (Kaznatcheev, 2010) would fare as the environment turns hostile. Will groups overcome their biases against each other, or will they compete even more for the more limited resource? Nearly 150 years after Peter Kropotkin’s Siberian expedition, the curtain is still up and basic questions on mutual aid in austere environments remain!
References
Hauert, C., Holmes, M., & Doebeli, M. (2006). Evolutionary games and population dynamics: maintenance of cooperation in public goods games. Proceedings of the Royal Society B: Biological Sciences, 273(1600): 2565-2571
Kaznatcheev, A. (2010). Robustness of ethnocentrism to changes in inter-personal interactions. Complex Adaptive Systems – AAAI Fall Symposium. [pdf]
Melbinger, A., Cremer, J., & Frey, E. (2010). Evolutionary game theory in growing populations. Physical Review Letters, 105(17): 178101. [arXiv pdf]
Shultz, T. R., Hartshorn, M., & Kaznatcheev, A. (2009). Why is ethnocentrism more common than humanitarianism? In N. A. Taatgen & H. van Rijn (Eds.), Proceedings of the 31st Annual Conference of the Cognitive Science Society (pp. 2100-2105). Austin, TX: Cognitive Science Society. [pdf]
Strassmann, J., & Queller, D. (2011). Evolution of cooperation and control of cheating in a social microbe Proceedings of the National Academy of Sciences, 108 (2), 10855-10862 DOI: 10.1073/pnas.1102451108


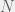 individuals and
individuals and 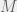 sets, where each individual can belong to
sets, where each individual can belong to  of the sets. Strategy and set membership are heritable (with mutation probabilities
of the sets. Strategy and set membership are heritable (with mutation probabilities  and
and  , respectively), and interactions are only with agents that share a set (if two agents share more than one set then they interact more than once). However, reproduction is inviscid: a random individual is selected to die and everybody competes to replace them with a child. This set-dependent interaction, makes the model equivalent to the
, respectively), and interactions are only with agents that share a set (if two agents share more than one set then they interact more than once). However, reproduction is inviscid: a random individual is selected to die and everybody competes to replace them with a child. This set-dependent interaction, makes the model equivalent to the 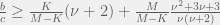
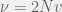 is the population-scaled set mutation rate, even when this is zero we get cooperation when
is the population-scaled set mutation rate, even when this is zero we get cooperation when  . Alternatively, to simplify we can take the limit of
. Alternatively, to simplify we can take the limit of 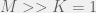 to get:
to get:
 ) and take the further limit of large populations
) and take the further limit of large populations 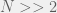 then this becomes a very simple:
then this becomes a very simple: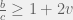
 of:
of:
 is a structural constant given by:
is a structural constant given by:
 ), and
), and  becomes:
becomes:
 , it is more enlightening to look at the reciprocal that in the limit of large
, it is more enlightening to look at the reciprocal that in the limit of large  becomes:
becomes:


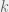 with categorization, we can study how much ethnocentric (and traitorous) agents are willing to pay for their greater cognitive abilities. This cost directly changes the default probability to reproduce (
with categorization, we can study how much ethnocentric (and traitorous) agents are willing to pay for their greater cognitive abilities. This cost directly changes the default probability to reproduce ( ), with humanitarians and selfish agents having
), with humanitarians and selfish agents having  and ethnocentrics and traitorous agents having
and ethnocentrics and traitorous agents having  . During each cycle, the
. During each cycle, the  and providing a benefit
and providing a benefit  (that varies depending on the simulation parameters) to the partner. For more detailed presentation of the simulation and default parameter, or just to follow along on your computer,
(that varies depending on the simulation parameters) to the partner. For more detailed presentation of the simulation and default parameter, or just to follow along on your computer, 
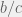 ratio and increasing
ratio and increasing 
 . From this limited data, I concluded that since the phase transition happens for
. From this limited data, I concluded that since the phase transition happens for 
 fixed at 0.01. Unsurprisingly, the transitions for proportion of ethnocentrics and humanitarians are indistinguishable, but without a proper analysis it is not clear if the transition from high to low cooperation also always coincides. For
fixed at 0.01. Unsurprisingly, the transitions for proportion of ethnocentrics and humanitarians are indistinguishable, but without a proper analysis it is not clear if the transition from high to low cooperation also always coincides. For  , agents are willing to invest more than
, agents are willing to invest more than 

 — a rather competitive environment. Matching Shultz, Hartshorn, & Kaznatcheev (2009) and consistent with Milbiner, Cremer, & Frey (2010), we can see an early spike in the number of cooperators as the world reaches its carrying capacity. After this transient period, the dynamics shift and defection becomes more competitive. The dynamics settle to a stable distribution of cooperators and defectors. The proportion of cooperation depends heavily on the environmental austerity. In a harsh environment with a low default birth rate of 0.24, the agents band together and cooperate and in a plentiful environment with high default birth rate of 0.28, defection dominates. As Kropotkin observed: cooperation is essential to surviving environmental austerity.
— a rather competitive environment. Matching Shultz, Hartshorn, & Kaznatcheev (2009) and consistent with Milbiner, Cremer, & Frey (2010), we can see an early spike in the number of cooperators as the world reaches its carrying capacity. After this transient period, the dynamics shift and defection becomes more competitive. The dynamics settle to a stable distribution of cooperators and defectors. The proportion of cooperation depends heavily on the environmental austerity. In a harsh environment with a low default birth rate of 0.24, the agents band together and cooperate and in a plentiful environment with high default birth rate of 0.28, defection dominates. As Kropotkin observed: cooperation is essential to surviving environmental austerity.


Diversity and persistence of group tags under replicator dynamics
November 29, 2015 by Artem Kaznatcheev 3 Comments
Everyday I walk to the Stabile Research Building to drink espresso and sit in my cozy — although oversaturated with screens — office. Oh, and to chat about research with great people like Arturo Araujo, David Basanta, Jill Gallaher, Jacob Scott, Robert Vander Velde and other Moffitters. This walk to the office takes about 30 minutes each way, so I spend it listening to podcasts. For the past few weeks, upon recommendation from a friend, I’ve started listing to the archive of the Very Bad Wizards. This is a casual — although oversaturated with rude jokes — conversation between David Pizarro and Tamler Sommers on various aspects of the psychology and philosophy of morality. They aim at an atmosphere of two researchers chatting at the bar; although their conversation is over Skype and drinks. It is similar to the atmosphere that I want to promote here at TheEGG. Except they are funny.
While walking this Wednesday, I listed to episode 39 of Very Bad Wizards. Here the duo opens with a Wilson & Haidt’s TIME quiz meant to quantify to what extent you are liberal or conservative.[1] They are 63% liberal.[2]
To do the quiz, you are asked to rate 12 statements (well, 11 and one question about browsers) on a six point Likert scale from strongly disagree to strongly agree. Here are the three that caught my attention:
Do you strongly agree? Strongly disagree? What was your overall place on the liberal-conservative scale?
Regardless of your answers, the statements probably remind you of an important aspect of your daily experience. The world is divided into a diversity of groups, and they coexist in a tension between their arbitrary, often artificial, nature and the important meaning that they hold to both their own members and others. Often this division is accompanied by ethnocentrism — a favoring of the in-group at the expensive of, or sometimes with direct hostility toward, the out-group — that seems difficult to circumvent through simply expanding our moral in-group. These statements also confront you with the image of what a world without group lines might look like; would it be more cooperative or would it succumb to the egalitarian dilemma?[5]
As you know, dear reader, here at TheEGG we’ve grappled with some of these questions. Mostly by playing with the Hammond & Axelrod model of ethnocentrism (2006; also see: Hartshorn, Kaznatcheev & Shultz, 2012). Recently, Jansson’s (2015) extension of my early work on the robustness of ethnocentrism (Kaznatcheev, 2010) has motivated me to continue this thread. A couple of weeks ago I sketched how to reduce the dimensionality of the replicator equations governing tag-based games. Today, I will use this representation to look at how properties of the game affect the persistence and diversity of tags.
Read more of this post
Filed under Analytic, Commentary, Models, Preliminary, Technical Tagged with cognitive science, ethics and morality, ethnocentrism, evolution of cooperation, replicator dynamics, Robert Axelrod