Effective games from spatial structure
December 7, 2018 5 Comments
For the last week, I’ve been at the Institute Mittag-Leffler of the Royal Swedish Academy of Sciences for their program on mathematical biology. The institute is a series of apartments and a grand mathematical library located in the suburbs of Stockholm. And the program is a mostly unstructured atmosphere — with only about 4 hours of seminars over the whole week — aimed to bring like-minded researchers together. It has been a great opportunity to reconnect with old colleagues and meet some new ones.
During my time here, I’ve been thinking a lot about effective games and the effects of spatial structure. Discussions with Philip Gerlee were particularly helpful to reinvigorate my interest in this. As part of my reflection, I revisited the Ohtsuki-Nowak (2006) transform and wanted to use this post to share a cute observation about how space can create an effective game where there is no reductive game.

Suppose you were using our recent game assay to measure an effective game, and you got the above left graph for the fitness functions of your two types. On the x-axis, you have seeding proportion of type C and on the y-axis you have fitness. In cyan you have the measured fitness function for type C and in magenta, you have the fitness function for type D. The particular fitnesses scale of the y-axis is not super important, not even the x-intercept — I’ve chosen them purely for convenience. The only important aspect is that the cyan and magenta lines are parallel, with a positive slope, and the magenta above the cyan.
This is not a crazy result to get, compare it to the fitness functions for the Alectinib + CAF condition measured in Kaznatcheev et al. (2018) which is shown at right. There, cyan is parental and magenta is resistant. The two lines of best fit aren’t parallel, but they aren’t that far off.
How would you interpret this sort of graph? Is there a game-like interaction happening there?
Of course, this is a trick question that I give away by the title and set-up. The answer will depend on if you’re asking about effective or reductive games, and what you know about the population structure. And this is the cute observation that I want to highlight.


 and
and 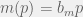 . After a long meeting with Robert a few month ago, I think that these were misleading payoffs to consider. I jotted these notes after the meeting, but forgot to release them on the blog. Instead, you get to enjoy them now while I refresh my memory.
. After a long meeting with Robert a few month ago, I think that these were misleading payoffs to consider. I jotted these notes after the meeting, but forgot to release them on the blog. Instead, you get to enjoy them now while I refresh my memory.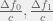 — the difference in the benefit of the public good from having 1 instead of 0 and 2 instead of 1 contributor in the group, respectively; measured in multiples of the cost c. In that case, if we want to recreate
— the difference in the benefit of the public good from having 1 instead of 0 and 2 instead of 1 contributor in the group, respectively; measured in multiples of the cost c. In that case, if we want to recreate  then just set
then just set  and
and  . Alternatively, if you want a free public good (c = 0) then use
. Alternatively, if you want a free public good (c = 0) then use  and
and  . I’ll leave verifying the arithmetic as an exercise for you, dear reader.
. I’ll leave verifying the arithmetic as an exercise for you, dear reader.


Software monocultures, imperialism, and weapons of math destruction
September 29, 2018 by Artem Kaznatcheev 7 Comments
This means that a bug that some engineers missed at Facebook compromised the security of users on completely unrelated sites like, say, StackExchange (SE) or Disqus — or any site that you can log into using your Facebook account.
A case of software monoculture — a nice metaphor I was introduced to by Jonathan Zittrain.
This could easily have knock-on effects for security. For example, I am one of the moderators for the Theoretical Computer Science SE and also the Psychology and Neuroscience SE. Due to this, I have the potential to access certain non-public information of SE users like their IP addresses and hidden contact details. I can also send communications that look much more official, along-side expected abilities like bans, suspensions, etc. Obviously, part of my responsibility as a moderator is to only use these abilities for proper reasons. But if I had used Facebook — disclosure: I don’t use Facebook — for my SE login then a potential hacker could get access to these abilities and then attempt phishing or other attacks even on SE users that don’t use Facebook.
In other words, the people in charge of security at SE have to worry not only about their own code but also Facebook (and Google, Yahoo!, and other OpenIDs).
Of course, Facebook is not necessarily the worst case of software monoculture or knock-on effects that security experts have to worry about. Exploits in operating systems, browsers, serves, and standard software packages (especially security ones) can be even more devastating to the software ecology.
And exploits of aspects of social media other that login can have more subtle effects than security.
The underlying issue is a lack of diversity in tools and platforms. A case of having all our eggs in one basket. Of minimizing individual risk — by using the best available or most convenient system — at the cost of increasing systemic risk — because everyone else uses the same system.
We see the same issues in human projects outside of software. Compare this to the explanations of the 2008 financial crises that focused on individual vs systemic risk.
But my favourite example is the banana.
In this post, I’ll to sketch the analogy between software monoculture and agricultural monoculture. In particular, I want to focus on a common element between the two domains: the scale of imperial corporations. It is this scale that turns mathematical models into weapons of math destructions. Finally, I’ll close with some questions on if this analogy can be turned into tool transfer: can ecology and evolution help us understand and manage software monoculture?
Read more of this post
Filed under Commentary Tagged with application of theory, current events, public good