Where did the love come from? Inclusive fitness vs. group selection
March 25, 2013 17 Comments
Altruism is widespread in the animal world, yet it seems to conflict with the picture of nature “red in tooth and claw” often associated with Evolution. One solution to this apparent paradox is to remember that the unit of selection is never the individual itself but the genes it carries. Thus, altruism may be explained if the altruist shares genes with the individual it helps in such a way that, while harming itself as an individual, it favors the spread of its genes. This idea of analyzing selection at the level of genes rather than the individual dates back to the 1930s, when Darwin’s theory and Mendelian genetics were first combined to form a unified framework now known as the neo-Darwinian synthesis.

Altruism is a common feature of animal behaviour. In this picture, a chimp mother helping another down a tree. Source: The Selfishness of Giving by Frans de Waal on Huffington Post.
Inclusive Fitness and Hamilton’s rule
But not until a few decades later, when D.W. Hamilton proposed his famous rule, was the idea integrated into evolutionary models. As Hamilton himself noted in his seminal 1964 paper: “With very few exceptions, the only parts of the theory of natural selection which have been supported by mathematical models admit no possibility of the evolution of any characters which are on average to the disadvantage of the individuals possessing them.” (Hamilton 1964) Early models focused on individual fitness without considering implications for relatives, with the notable exception of parental care. Filling this vacuum, Hamilton proposed a simple model which aimed to explain how self-harming social behavior could evolve. The basic idea is, again, to look at the effects of an action on the altruist’s “inclusive fitness” – i.e. on all the copies of its genes. To do this, one must add up the reproductive benefit (
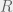


In formulating this rule, Hamilton did not intend to create a new theory distinct from natural selection. Rather, his model attempts to fully account for the genetic underpinning of natural selection. As has been amply noted in recent literature, much criticism of inclusive fitness theory (IFT) stems from the mistaken belief that inclusive fitness is distinct from “regular” natural selection. But before I describe further the controversy surrounding inclusive fitness, let us consider a competing, though not exclusive, explanation for the evolution of altruism: group selection.
Group selection: an alternative mechanism for the evolution of altruism
Group selection is the selection of altruistic traits resulting from the differential survival of entire groups of organisms. Three requirements must be met for group selection to happen: (1) the groups have to be reproductively isolated, at least to a degree; (2) the new altruistic trait must improve the chances of survival of the trait-carrying group relative to other groups; and (3) the gene that causes the trait must establishes itself first in a small group through genetic drift. The first two conditions are self-explanatory if we remember that the groups themselves must be the units of selection. As for the third, it can be understood with the use of an example borrowed from Maynard Smith. Consider (partly) reproductively isolated groups of Anubis baboons. Anubis baboons live in large troops; males must leave their troop to mate while females always remain in their native group. As a result, males are typically genetically far from other males, females, and all offspring beside their own, whereas females of the same troop are closely related. Note further that infanticide by unrelated adult male baboons is common. Suppose now that a mutation causes some females to protect the offspring of other females in their troop and that the mutant gene spreads through the group whose fitness increases as a result. Is group selection at play here? The example was chosen so that the first two conditions – reproductive isolation and differential extinction – are met. However, we have to explain how the altruistic trait spread through the group in the first place. Since the group is large, genetic drift (gene frequency changes due to random gene sampling at each generation) cannot be responsible for the gene’s spread. Instead, it is likely that the mutation increases the inclusive fitness of the altruistic female, giving it a selective advantage. Or, we can imagine that the apparently altruistic action in fact helps the defendant female through, say, reciprocal defense. Thus, when genetic drift does not the cause the trait’s initial spread, even if it eventually benefits the entire group, selection for altruism cannot be explained by group selection alone (Maynard Smith 1976).
Empirical tests
Inclusive fitness and group selection – at least as I understand them – both are logically consistent with natural selection and with each other. Yet, during the last decades, the two theories have split the scientific community into two warring camps, each holding tightly to a theory and denigrating the other, sometimes with rather “unscientific” ardor and partiality. The disagreement stems partly from a misunderstanding. In a recent paper highly critical of inclusive fitness, Nowak et al. argue that IFT is a superfluous construct , an “abstract entreprise” divorced from natural selection. They claim that inclusive fitness theory (1) only considers the direct effects of altruistic actions while ignoring all retroactive effects of the recipients on the actor; (2) ignores all synergistic effects and assumes that the effects of the action are additive; (3) can only be applied to special types of population structure, namely static structures (fixed interactions between individuals) and “0 or 1” dynamic structures (where individuals either interact or don’t) (Nowak et al. 2010). While particular models may be rightful targets of this criticism, inclusive fitness itself is not. These restrictions are features of (necessarily) simplified models – especially those in textbooks – not of the theory as such.
Group selection and kin selection both being compatible with natural selection, we can only arbitrate between them by comparing their empirical merits. One domain in which this has been done is the study of eusocial animals. Eusociality – a phenomenon observed primarily among insects- occurs when labor, defense and reproduction are divided among castes in a colony. Eusociality involves altruism since all workers and warriors sacrifice their direct reproductive potential to support the reproducing caste. Early inclusive fitness theorists posited that in colonies with non-overlapping generations, eusociality should be more prominent where sibling-relatedness is high because in that case the inclusive fitness benefits of altruism are greater. Some theorists predicted, for instance, that eusociality is more widespread in insects exhibiting haplodiploidy – a type of sex determination whereby unfertilized eggs give birth to males and fertilized eggs, to females. Since they inherit the integrity of their father’s genotype, haplodiploid sisters share a large portion of their genes. Hence, altruistic and eusocial behavior should be more likely to evolve in haplodiploids.
Or so theorists claimed, until data collected in the last decades proved them wrong: haplodiploidy and eusociality are not statistically correlated. Nowak et al. claimed that this failure invalidates IFT, whose predictive results, they say, have been meager. But in doing so, they once again threw the baby with the bathwater, so to speak (Nowak et al. 2010). Haplodiploidy aside, IFT explains a wide range of phenomena. Shortly after Nowak et al.’s attack on the IFT, 137 authors (!) wrote a scathing response that drew attention to IFT’s predictive and explanatory power. For instance, colonies with single queens are statistically more likely to be eusocial, probably for the same reason that haplodiploidy was expected to favor eusociality: in single-queen colonies siblings share on average a greater portion of their genotypes (Abbot et al. 2011). But why then is haplodiploidy, unlike single-queen colony structure, not correlated with eusociality? One possible reason is that the inclusive fitness benefits of altruism are overwhelmed by competition since, when siblings compete, the reproductive cost (
In short, group selection and inclusive fitness are both ways to account for the genetic basis of natural selection. As such, neither is a separate framework competing with classical evolution, and both may provide useful explanations and predictions in understanding altruism – as has been amply shown for inclusive fitness. More research is needed, however, to assess their relative importance in the evolution of altruistic behavior.
References
Abbot, P., Abe, J., Alcock, J., Alizon, S., Alpedrinha, J. A., Andersson, M., … & Gardner, A. (2011). Inclusive fitness theory and eusociality. Nature, 471(7339): E1-E4.
Hamilton, W. W. (1964). The genetical evolution of social behaviour 1. J. Theoret. Biol., 7: 1-16.
Maynard Smith, J. (1976). Group selection. Quarterly Review of Biology, 51(2): 277-283.
Mulder, M. B. (2007). Hamilton’s rule and kin competition: The kipsigis case. Evolution and Human Behavior, 28: 299-312.
Nowak M.A., Tarnita C.E. & Wilson E.O. (2010). The evolution of eusociality, Nature, 466 (7310) 1057-1062. DOI: 10.1038/nature09205


Awesome post, Eric! Thank you for writing it.
I have a hard time understanding why inclusive fitness should be considered a theory instead of a framework within evolution. Can you think of an example of some observation that would falsify inclusive fitness (the broad concept, not a specific model like the one Nowak, Tarnia, and Wilson attack) but not evolution as a whole? I have a hard time coming up with such a hypothetical experiment.
My personal qualm with IF, or at least with Hamilton’s rule, is that I feel people attribute too much to it. In spatial models for instance, for relatedness to follow Hamilton’s rule it needs to encode things like the organisms current position and other non-genetic factors… it seems to be a real stretch to call such a parameter ‘relatedness’ in any meaningful way, and it definitely doesn’t capture Hamilton’s original intention. If you let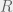 , and
, and  be as complicated as you want (note that b and c by themselves don’t make sense since they are measured in arbitrary units, only their ratio can make sense) then the theory looses any real scientific power, and becomes a trivial observation about linear systems (any transition in a linear system will necessarily be of the form
be as complicated as you want (note that b and c by themselves don’t make sense since they are measured in arbitrary units, only their ratio can make sense) then the theory looses any real scientific power, and becomes a trivial observation about linear systems (any transition in a linear system will necessarily be of the form 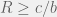 for some sufficiently complicated
for some sufficiently complicated 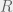 and
and  ).
).
Of course, group selection also suffers from the often arbitrary distinction between what constitutes a group and how groups “reproduce”. However, it can still often be a powerful well-grounded tool, for instance if your group is “cell” and your individuals are “plasmids”:
Paulsson, J. (2002). Multileveled selection on plasmid replication. Genetics, 161(4): 1373-1384.
Very interesting post. I found Pinker’s essay on group selection immensely clarifying: http://edge.org/conversation/the-false-allure-of-group-selection. In particular, for group selection to be important, the same preconditions must apply as for evolution by natural selection: groups must reproduce themselves, groups must have differential fitness (some groups must reproduce more effectively than others, or some groups must go extinct), and group properties must be heritable. There’s a huge timescale problem here, since it is very hard to imagine scenarios where groups reproduce quickly enough to override the selective effects of a trait that is disadvantageous at the IF level. I was on the fence about group selection for a long time, but Pinker’s essay convinced me that it is highly unlikely to have played a significant role in evolution. (But now I have to go look at the article Artem recommended :->).
Artem, your comment on spatiality is interesting. But if spatial position *correlates* with relatedness, then altruism towards spatially proximate individuals is explained by IF, isn’t it? IF does not demand that organisms actually *know* who is related to them and who is not. It only demands that they be able to act in such a manner as to favor those who are related to them, on average. If spatial proximity can act as a proxy for relatedness, that still seems like IF to me.
Thanks for the comments guys.
Regarding the limits of group selection. As Ben observed, the typically large generation time of groups along with several other limitations (the need for spread through genetic drift, “cheaters” sneaking into the group and abusing altruism) make it difficult to imagine how group selection -when defined as “natural selection on groups”- could have been a major force in evolution.
But the paper Artem recommended seems to give an example that fits with these requirements, if we define the procaryotic cell as the group and the plasmids it contains as the individuals. There the generation time of the individuals and the group are at least comparable (I believe…). And it’s true that the definition of “group” is problematic because it’s arbitrary, but so is, ultimately, any definition of the “individual”. For instance in coral colonies exhibiting organism-like division of labor it’s hard to pinpoint where the individual ends and where the group begins.
However, to be fair, the stringent definition of group selection I gave in my post is not the one commonly used by all group selectionists. The definition of group selection seems to have recently shifted from “natural selection on groups” – where the groups are the units of selection – to “natural selection divided into within-group and without- group components” . In the post I omitted the latter definition for the sake of brevity, and also because I feel that this definition deprives “group selection” of its original meaning (in addition to making it formally indistinguishable from IFT and hence redundant. See Marshall 2011: “Group selection and kin selection: formally equivalent approaches”)
Ben, it isn’t just “correlates” it is “part of the definition”. I.e. if you take the same exact agent (genetically) and place him in a different location, he will have a different relatedness to the focal individual. Hence, it isn’t just a proxy, it is taking a quantity and calling it “relatedness” when it doesn’t correspond in any intuitive way to the word.
Integrating position into relatedness does seem like a misuse of the term.
However, it seems to me like IF can be applied to spatial model while keeping the usual definition of relatedness as pedigree or as some other form of genetic association (e.g. the “greenbeard” effect). Position can be used to determine costs and benefits of particular actions with c and b depending on relative position of actor/recipient. Or am I missing something?
In the first version of this post I gave the wrong definition of haplodiploidy by saying that unfertilized eggs give birth to females in haplodiploid species. In fact, it is the males who are haploid whereas the males are diploid. The mistake has been corrected, sorry to the first readers!
oops. I meant: “In fact, it is the males who are haploid whereas the females are diploid”
Pingback: Continuing our exploration of group selection | Theory, Evolution, and Games Group
Pingback: Evolutionary economics and game theory | Theory, Evolution, and Games Group
Pingback: Social learning dilemma | Theory, Evolution, and Games Group
Pingback: Evolutionary games in set structured populations | Theory, Evolution, and Games Group
Pingback: Stats 101: an update on readership | Theory, Evolution, and Games Group
Pingback: Liquidity hoarding and systemic failure in the ecology of banks | Theory, Evolution, and Games Group
Pingback: Cataloging a year of blogging: applications of evolutionary game theory | Theory, Evolution, and Games Group
Pingback: Computational theories of evolution | Theory, Evolution, and Games Group
Pingback: Operationalizing replicator dynamics and partitioning fitness functions | Theory, Evolution, and Games Group
Pingback: Short history of iterated prisoner’s dilemma tournaments | Theory, Evolution, and Games Group