Ohtsuki-Nowak transform for replicator dynamics on random graphs
October 25, 2012 21 Comments
We have seen that the replicator equation can be a useful tool in understanding evolutionary games. We’ve already used it for a first look at perception and deception, and the cognitive cost of agency. However, the replicator equation comes with a number of inherent assumptions and limitations. The limitation Hisashi Ohtsuki and Martin Nowak wanted to address in their 2006 paper “The replicator equation on graphs” is the assumption of a totally inviscid population. The goal is to introduce a morsel of spatial structure, while still retaining the analytic tractability and easy-of-use of replicator dynamics.
Replicator dynamics assume that every individual in population is equally likely to interact with every other individual in the population. However, in most settings the possible interactions are not as simple. Usually, there is some sort of structure to the population. The way we usually model structure, is by representing individuals as vertexes in a graph, and if two vertexes are adjacent (connected by an edge) then the two individuals can interact. For replicator dynamics the model would be a complete graphs. Ohtsuki and Nowak consider a structure which is a bit more complicated: a random 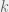
There are several ways to think about random
With the spatial structure in place, the next important decision is the update or evolutionary dynamics to use. Ohtsuki & Nowak (2006) consider four different rules: birth-death (BD), death-birth (DB), imitation (IM), and pairwise comparison (PC). For BD, player is chosen for reproduction in proportion to fitness and the offspring replaces a neighbor chosen uniformly at random. In DB, a focal agent is chosen uniformly at random and is replaced by one of his neighbors, who are sampled randomly in proportion to fitness. IM is the same as DB, except the focal agents counts as one of its own neighbors. For PC, both the focal individual and his partner are chosen uniformly at random, but the focal agent replaces his partner with probability that depends (according to the Fermi rule) on their payoff difference, otherwise the partner replaces the focal individual.
Throughout the paper, fitness is calculated in the limit of weak selection. To account for the spatial structure, the authors use pair-approximation. This method is perfect for Bethe lattice and accurate for any graph that is locally tree like. Thankfully, random graphs are locally tree like (out to distance on the order of 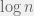

The stunning aspect of the paper, is that in the limit of very large populations they get an exceptionally simple rule. Give a game matrix 
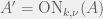
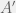
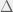
![A = [a_{ij}]](https://s0.wp.com/latex.php?latex=A+%3D+%5Ba_%7Bij%7D%5D&bg=f0f0f0&fg=555555&s=0&c=20201002)
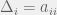
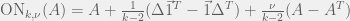
Where 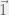

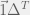

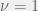
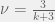
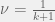

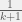
My other reason for rewriting in this condensed form is to gain an intuition for each term of the sum. The first term is the original game, and is expected to be there with some summation because we are doing a small perturbation in the limit of weak selection, thus a linear correction to 
As case studies, Ohtsuki & Nowak (2006) apply their transform to the prisoner’s dilemma, snowdrift, coordination, and rock-paper scissors games. I refer the interested reader to the original paper for the latter example and I will deal with the former three at once by looking at all cooperate-defect games:

Where X-Y is a new coordinate system given by the linear transform:


The effects of this coordinate transform on UV-space can be seen in the figure above for 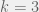
![]()
Hilbe, C. (2011). Local replicator dynamics: a simple link between deterministic and stochastic models of evolutionary game theory. Bull Math Biol, 73: 2068-2097.
Ohtsuki H, & Nowak MA (2006). The replicator equation on graphs. Journal of Theoretical Biology, 243 (1), 86-97 PMID: 16860343


Would other update rules fall in between those two in UV space, or might there be even wilder changes due to update rule?
IM falls in between them, as I suspect would the probabilistic rule I suggest. However, I don’t think you can reasonable take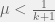 without making odd update rules.
without making odd update rules.
As far as I can tell, that term is related to the number of agents you sample to get the most important info for an update. It is unusual to look more than two steps away from a focal individual to calculate where his child goes, and the number of agents within two steps is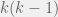 and the finite-sampling discount factor according to Hilbe would go as
and the finite-sampling discount factor according to Hilbe would go as 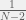 , so having more than
, so having more than 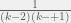 of a discount seems unlike to me and DB achieves this.
of a discount seems unlike to me and DB achieves this.
However, I will think about it more carefully and try to expand on the ideas when I post about Hilbe's paper. Thanks for the suggestion!
Pingback: Social learning dilemma | Theory, Evolution, and Games Group
Pingback: Evolutionary games in set structured populations | Theory, Evolution, and Games Group
Pingback: Natural algorithms and the sciences | Theory, Evolution, and Games Group
Pingback: Game theoretic analysis of motility in cancer metastasis | Theory, Evolution, and Games Group
Pingback: Microenvironmental effects in prostate cancer dynamics | Theory, Evolution, and Games Group
Pingback: Edge effects on the invasiveness of solid tumours | Theory, Evolution, and Games Group
Pingback: Computational complexity of evolutionary stable strategies | Theory, Evolution, and Games Group
Pingback: Cooperation through useful delusions: quasi-magical thinking and subjective utility | Theory, Evolution, and Games Group
Pingback: Approximating spatial structure with the Ohtsuki-Nowak transform | Theory, Evolution, and Games Group
Pingback: Cataloging a year of blogging: cancer and biology | Theory, Evolution, and Games Group
Pingback: Space and stochasticity in evolutionary games | Theory, Evolution, and Games Group
Pingback: Seeing edge effects in tumour histology | Theory, Evolution, and Games Group
Pingback: Making model assumptions clear | CancerEvo
Pingback: Measuring games in the Petri dish | Theory, Evolution, and Games Group
Pingback: Spatializing the Go-vs-Grow game with the Ohtsuki-Nowak transform | Theory, Evolution, and Games Group
Pingback: Token vs type fitness and abstraction in evolutionary biology | Theory, Evolution, and Games Group
Pingback: Abstract is not the opposite of empirical: case of the game assay | Theory, Evolution, and Games Group
Pingback: Heuristic models as inspiration-for and falsifiers-of abstractions | Theory, Evolution, and Games Group
Pingback: Effective games from spatial structure | Theory, Evolution, and Games Group