Evolutionary games in set structured populations
May 8, 2013 3 Comments
We have previously discussed the importance of population structure in evolutionary game theory, and looked at the Ohtsuki-Nowak transform for analytic studies of games on one of the simplest structures — random regular graphs. However, there is another extremely simple structure to consider: a family of inviscid sets. We can think of each agent as belonging to one or more sets and interacting with everybody that shares a set with them. If there is only one set then we are back to the case on a completely inviscid population. If we associate a set with each edge in a graph and restrict them to have constant size then we have standard evolutionary graph theory. However, it is more natural to allow sets to grow larger if their members have high fitness.
Tarnita et al. (2009) consider a population of 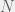
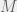



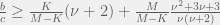
Where 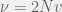

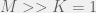

If we allow the maximum number of sets (
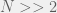
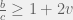
Finally, in the supplementary materials, the authors derive a very nice relationship for evolution of cooperation on arbitrary cooperate-defect games given by a payoff matrix 

where 

Which in the limit of small population-scaled strategy mutation (


Since the structural constant is always greater than 1, and since we typically care about large 


And simplifies the general equation for cooperation to emerge to:

Which can be seen as a relaxation of the classic game theory concept of risk-dominance.
![]() Tarnita, C., Antal, T., Ohtsuki, H., & Nowak, M. (2009). Evolutionary dynamics in set structured populations Proceedings of the National Academy of Sciences, 106 (21), 8601-8604 DOI: 10.1073/pnas.0903019106
Tarnita, C., Antal, T., Ohtsuki, H., & Nowak, M. (2009). Evolutionary dynamics in set structured populations Proceedings of the National Academy of Sciences, 106 (21), 8601-8604 DOI: 10.1073/pnas.0903019106

